TSS seçimi
aktiv sintetik promoterlərin yaradılması - Süni intellekt insan genlərinin idarəetmə - Transkripsiya başlanğıc yerləri (TSS) seçimi əvvəllər təsvir edildiyi kimi aparılmışdır. Prosesin qısa xülasəsi belədir: biz GENCODE tərəfindən müəyyən edilmiş TSS-ləri seçdik və əlavə şərt olaraq onların FANTOM5 məlumat bazasına əsasən ən azı bir hüceyrə növündə və ya toxumada aktiv olmasını tələb etdik. Bu proses nəticəsində 30,607 TSS-dən ibarət dəqiqləşdirilmiş bir dəst yaradıldı.
Genom miqyaslı MPRA məlumat dəsti
HepG2 və K562 hüceyrələri üçün ilkin PARM modelləri, ictimaiyyətə açıq olan genom miqyaslı MPRA məlumatları (GEO giriş nömrəsi GSE128325) ilə təlim edilmişdir. Biz bütün kitabxanalardan fraqmentləri birləşdirdik və 30,607 TSS-ə nisbətən -300 bp-dən +100 bp-ə qədər olan pəncərə ilə üst-üstə düşənləri seçdik.
MPRA kitabxanalarının yaradılması
Hədəflənmiş kitabxananın DNT fraqmentasiyası və tutma üsulları ilə hazırlanması
Hədəflənmiş kitabxananı yaratmaq üçün insan hüceyrə xəttindən (HG02601) "Isolate II Genomic DNA kit" (Bioline, BIO-52066) dəsti ilə 100 µq DNT izolyasiya edildi. İzolyasiya edilmiş DNT daha sonra "dsDNA Fragmentase" (NEBNext, M0348L) vasitəsilə 30 dəqiqə ərzində fraqmentlərə ayrıldı və təxminən 200-400 bp ölçüdə olan fraqmentlər gel ekstraksiyası ilə seçildi. DNT-nin ucları "End-IT DNA End-Repair kit" (Lucigen, ER81050) dəsti ilə bərpa edildi və ardınca "Klenow Fragment (3′→5′ Exo-; NEB, M0212M)" istifadə edilərək A-quyruq əlavə edildi. Klonlaşdırma məqsədilə, fraqmentlərin 5′ və 3′ ucları üçün T-çıxıntısı olan iki xüsusi 31 bp-lik dsDNA adapteri (oNK46 və oNK47), "Quick Ligation kit" (NEB, M2200L) ilə TA-liqasiyası vasitəsilə fraqmentlərə birləşdirildi (oliqonukleotid ardıcıllıqları üçün Əlavə Cədvəl 1-ə baxın). Bu adapterlər, hibridləşmədən sonra fraqmentlərin Gibson birləşməsinə imkan vermək üçün xətti barkodlu p101 vektorunun 3′ və 5′ ucları ilə üst-üstə düşür. PZR gücləndirilməsi oNK51 və oNK52 primerləri və "Equinox" polimerazası (Twist, 104176) ilə səkkiz tsikl ərzində aparıldı. TSS-in promotor bölgəsini tutmaq üçün biz 30,607 TSS və onların -300 ilə +100 bp pəncərəsini seçdik və bu xüsusi bölgələr üçün Twist şirkətindən 127,575 zonddan ibarət yüksək dəqiqlikli hibridləşmə tutma kitabxanası sifariş etdik. Zondların fraqmentlərimizə qeyri-spesifik bağlanmasının qarşısını almaq üçün xüsusi Gibson adapterlərinə (oNK57 və oNK58) komplementar olan xüsusi blokerlər hazırladıq. Fraqmentlər istehsalçının protokollarına uyğun olaraq xüsusi hibridləşmə paneli ilə tutuldu. Proses zamanı 1 µq fraqment, spesifik (xüsusi dizayn edilmiş) və qeyri-spesifik (COT-1 DNT) bloker məhlulunun iştirakı ilə 16 saat ərzində xüsusi hibridləşmə paneli ilə hibridləşdirildi. Sonra, hibridləşmiş fraqmentlər streptavidin bağlanması ilə zənginləşdirildi və oNK51 və oNK52 primerləri ilə doqquz tsikllik PZR vasitəsilə gücləndirildi. Tutulmuş fraqmentlər təmizləndi və "HiFi DNA assembly master mix" (NEB, E2621L) ilə 50 °C-də 60 dəqiqə ərzində Gibson birləşməsi vasitəsilə xətti barkodlu p101 vektoruna klonlaşdırıldı. Gibson birləşməsi qarışığı təmizləndi və istehsalçının protokollarına uyğun olaraq "MegaX DH10B" ultrakompetent bakteriyalarına (Thermo, C640003) transformasiya edildi. Transformasiya edilmiş bakteriyalar bir gecə LB mühitində böyüdüldü və plazmid kitabxanası "Gigaprep Isolation kit" (Thermo, K210009XP) dəsti ilə izolyasiya edildi.
Oliqonukleotid əsaslı kitabxanalar
Hər üç sintetik kitabxana (sintetik promotorlar, ISM və motiv daxiletmələri) Twist şirkətindən sifariş edilmiş sintetik oliqonukleotidlər vasitəsilə yaradıldı. Oliqonukleotid kitabxanası Twist-dən sifariş edildi və "KAPA HiFi Hotstart Readymix" (Roche, KK2601) istifadə edilərək oNK69 və oNK71 ilə 14 tsikl ərzində gücləndirildi. Kitabxana muncuqla təmizləndi, "End-IT DNA End-Repair kit" (Lucigen, ER81050) ilə ucları bərpa edildi, yenidən muncuqla təmizləndi və EcoRI-HF (NEB, R3101S) və NheI-HF (NEB, R3131S) ilə 37 °C-də 1 saat ərzində həzm edildi. Fraqmentlər, T4 DNT ligazası (Roche, 10799009001) istifadə edilərək NheI-HF/EcoRI-HF ilə ikiqat həzm edilmiş vektora (p101 vektorunun bir uyğunlaşması) liqasiya edildi. Liqasiya qarışığı yuxarıda təsvir edildiyi kimi təmizləndi və transformasiya edildi.
Hüceyrə kulturası və transfeksiya
K562 (ATCC-CCL 243), U2OS (ATCC HTB-96), HCT116 (ATCC CCL-247), LNCaP (ATCC CRL-1740), HepG2 (ATCC HB-8065), HEK293 (ATCC CRL-1573), MCF7 (ATCC HTB-22) və AGS (ATCC CRL-1739) hüceyrə xətlərinin hamısı Amerika Tip Mədəniyyət Kolleksiyasından (ATCC) əldə edilmişdir. HAP1 hüceyrə xətti isə bu hüceyrə xəttini yaratmış T. Brummelkamp-dan alınmışdır. Mənbələrin təsdiqlənmiş olduğunu nəzərə alaraq, hüceyrə xətləri yenidən təsdiqlənmədi. Bütün hüceyrə kulturaları mütəmadi olaraq mikoplazma üçün yoxlanıldı. K562 hüceyrələri RPMI1640-da, HepG2, MCF7 və HEK293 DMEM-də, HCT116 və U2OS McCoy5a-da, LNCaP və AGS RPMI1640-da, HAP1 isə 1% qlutamin və 25 mM HEPES ilə IMDM-də kultivasiya edildi. Bütün mühitlərə 10% FBS (Gibco) və 1% penisilin-streptomisin (Thermo) əlavə edildi. Bütün hüceyrələr əvvəllər təsvir edildiyi kimi transfeksiya edildi, lakin HCT116, LNCaP və MCF7 müvafiq olaraq D-032, T-009 və P-020 nukleofektor proqramları ilə, HEK293 və U2OS isə Lipofectamine 2000 və ya 3000 ilə transfeksiya edildi. HAP1 isə Xfect reagenti (Takara, PT5003-2) ilə transfeksiya olundu. Hədəflənmiş kitabxananın hər bir bioloji təkrarı üçün 10 milyon hüceyrə, K562 hüceyrələrindən istifadə edilən təcrübədə isə 50 milyon hüceyrə istifadə edildi. Hər hansı bir oliqonukleotid əsaslı kitabxananın hər bir bioloji təkrarı üçün 5 milyon hüceyrə istifadə edildi.
Orqanoid kulturası
CRC orqanoidi cərrahi rezeksiyadan əldə edilmiş və əvvəllər təsvir edildiyi kimi yaradılmışdır. Nümunə götürülməsi Niderland Xərçəng İnstitutu – Antoni van Leeuwenhoek Xəstəxanasının Tibbi Etika Komitəsi tərəfindən təsdiqlənmiş (NL48824.031.14) və xəstədən yazılı razılıq alınmışdır. Bu layihə üçün insan materialı və ya məlumatlarının istifadəsi NKI-AVL İnstitusional Nəzarət Şurası tərəfindən təsdiqlənmiş və IRBdm23-361 nömrəsi ilə qeydiyyata alınmışdır. Şiş toxuması mexaniki olaraq parçalandı və 1.5 mg ml⁻¹ kollagenaza II (Sigma-Aldrich), 10 μg ml⁻¹ hialuronidaza tip IV (Sigma-Aldrich) və 10 μM Y-27632 (Sigma-Aldrich) ilə həzm edildi. Hüceyrələr Cultrex RGF BME tip 2-yə (3533-005-02, R&D Systems) əkildi və CRC tam orqanoid mühiti əlavə edilməzdən əvvəl 20 dəqiqə 37 °C inkubatorda saxlanıldı. Mühit əvvəllər təsvir edildiyi kimi istifadə edildi və tərkibi Ad-DF+++ (Advanced DMEM/F12, Gibco), 2 mM Ultraglutamine I (Lonza), 10 mM HEPES (Gibco), hər biri 100 U ml⁻¹ olan penisilin və streptomisin (Gibco), 10% noggin-şərtləndirilmiş mühit, 20% R-spondin1-şərtləndirilmiş mühit, 1× B27 əlavəsi (A vitaminsiz, Gibco), 1.25 mM N-asetilsistein (Sigma-Aldrich), 10 mM nikotinamid (Sigma-Aldrich), 50 ng ml⁻¹ insan rekombinant EGF (Peprotech), 500 nM A83-01 (Tocris), 3 μM SB202190 (Cayman Chemicals) və 10 nM prostaglandin E₂-dən (Cayman Chemicals) ibarətdir. Orqanoidlər hər həftə TrypLE Express (Gibco) ilə fermentativ parçalanma yolu ilə pasaj edildi, yuyuldu və BME-yə yerləşdirildi. Geniş miqyaslı orqanoid artımı üçün əvvəllər təsvir edilmiş protokoldan istifadə edildi. Orqanoidlər SNP massivi ilə təsdiqləndi və Mycoplasma PCR43 və MycoAlert Mycoplasma Detection kit (LT07-318) ilə aylıq olaraq mikoplazma üçün yoxlanıldı.
Orqanoid elektroporasiyası
Elektroporasiya protokolu əvvəlki tədqiqatdan uyğunlaşdırılmışdır. Orqanoidlər normal kultura mühitində böyüdüldü və elektroporasiyadan 2 gün əvvəl təzəcə bölündü. Elektroporasiya günündə orqanoidlər TrypLE Express (Gibco) ilə tək hüceyrələrə fermentativ olaraq parçalandı və 5 dəqiqə ərzində 10 μM Y-27632 (Sigma-Aldrich) ilə əlavə edildi. Hüceyrələr otaq temperaturunda PBS (Gibco) ilə toplandı və 500g-də 5 dəqiqə fırladıldı. Hər reaksiya üçün 5 milyon hüceyrə istifadə edildi və 22 μg hədəflənmiş kitabxana ilə əlavə edilmiş 100 μl otaq temperaturunda Opti-MEM-də (ThermoFisher) resuspensiya edildi. Hər bir bioloji təkrar üçün ümumilikdə 50 milyon hüceyrə istifadə olundu. Elektroporasiya NEPA21 sistemi ilə aşağıdakı parametrlərlə aparıldı: porinq impuls dəyərləri gərginlik = 175 V, impuls uzunluğu = 5.0 ms, impuls intervalı = 50.0 ms, impuls sayı = 2, sönmə dərəcəsi = 10% və polyarlıq = +; və ötürmə impuls dəyərləri gərginlik = 20 V, impuls uzunluğu = 50 ms, impuls intervalı = 50 ms, impuls sayı = 5, sönmə dərəcəsi = 40% və polyarlıq = +/–. Elektroporasiya 2.0 mm aralıqlı küvetlərdə (Cell Projects) həyata keçirildi. Elektroporasiyadan sonra üzərinə 400 μl 10 μM Y-27632 ilə əlavə edilmiş Opti-MEM əlavə edildi və otaq temperaturunda 30 dəqiqə inkubasiya edildi. Daha sonra hüceyrələr plastik birdəfəlik pipetlə toplandı, 500g-də 5 dəqiqə fırladıldı və 5% BME ilə normal kultura mühitində (10 μM Y-27632 ilə əlavə edilmiş) resuspensiya edildi.
Şərait və təsirlər
İstilik şoku şəraiti üçün hüceyrələr kitabxana ilə transfeksiya edildi və 24 saat 37 °C-lik inkubatora yerləşdirildi. Toplanmadan üç saat əvvəl hüceyrələr 43 °C-yə ayarlanmış inkubatora köçürüldü və 3 saat sonra RNT izolyasiyası üçün toplandı. Kimyəvi təsirlər üçün həm nutlin-3a (MedChemExpess, HY-10029), həm də PMA (ThermoFisher, J63916.MCR) DMSO-da həll edildi və müvafiq olaraq 8 µM və 1 µM konsentrasiyalarda istifadə edildi. Birləşmələr transfeksiyadan dərhal sonra əlavə edildi və RNT izolyasiyası üçün 24 saat sonra toplandı.
Kitabxananın sekvenslənməsi
Hədəflənmiş kitabxananın xarakterizasiyası (barkod-fraqment əlaqəsi) əvvəllər təsvir edildiyi kimi iPCR ilə aparıldı. Kitabxana girişi (pDNA) də əvvəllər təsvir edildiyi kimi yaradıldı, lakin plazmid kitabxanasını həzm etmək üçün NcoI-HF istifadə edildi.
RNT nümunəsinin emalı
RNT TRIzol (ThermoFisher, 15596018) ilə izolyasiya edildi və hər reaksiyada 2.5 µg RNT olan 10 µl-lik reaksiyalara bölündü. Hər bioloji təkrar üçün paralel olaraq 12 reaksiya aparıldı. Hər reaksiyaya 60 dəqiqə ərzində 10 vahid DNase I (Roche, 04716728001) əlavə edildi və sonra 1 µl 25 mM EDTA əlavə edilərək 70 °C-də 10 dəqiqə saxlanmaqla inaktivləşdirildi. cDNT, GFP ORF-ni hədəfləyən gen-spesifik primerin (oNK17) 1 µl-i və dNTP-nin (hər biri 10 mM) 1 µl-i əlavə edilərək 65 °C-də 8 dəqiqə inkubasiya edilməklə yaradıldı. Bu reaksiya 4 µl RT buferi, 20 vahid RNase inhibitoru (Thermo, EO0381), 20 vahid Maxima tərs transkriptaza (Thermo, EP0743) və 2.5 µl su əlavə edilərək 50 °C-də 60 dəqiqə davam etdirildi və sonra 85 °C-də 5 dəqiqə istiliklə inaktivləşdirildi. Hər reaksiya MyTaq Red mix (Bioline, BIO-25043) ilə (indeks variantları) MMA219 və oNK28 istifadə edilərək PZR ilə gücləndirildi. PZR reaksiyaları muncuqla təmizləndi (GC Biotech, CPCR-0500) və Illumina NextSeq və ya NovaSeq 6000 platformalarında tək oxunuşlu sekvenslənməyə məruz qaldı.
Oxunuşların emalı
Sekvens oxunuşları əvvəllər təsvir edildiyi kimi emal edildi. Bu proses həm cDNT (fraqment aktivliyi), həm də iPCR və ya pDNA (giriş, normallaşdırma) üçün hər bir fərdi fraqment başına oxunuş saylarını göstərən bir cədvəllə nəticələndi. Xam dəyərləri milyonda oxunuşlara (RPM) çevirdikdən sonra, oxunuş saylarını aşağıdakı kimi normallaşdırdıq:
Nisbətfraqment,təkrar = (cDNT RPMfraqment,təkrar / iPCR RPM) + Q01(nisbəttəkrar)
Burada Q01(nisbəttəkrar) həmin təkrar üçün sıfırdan fərqli müşahidə olunan nisbətlərin 0.1 kvantilidir. Sonra, fraqmentin promoter fəallığı aşağıdakı kimi təyin edildi:
Fraqment aktivliyi = (ΣNtəkrar=1 log₂(nisbətfraqment)) / N
Burada N bioloji təkrarların ümumi sayıdır.
Modelin giriş məlumatları və promoter fəallığı
PARM, kitabxanadan tək bir fraqmentin genomik ardıcıllığını giriş olaraq qəbul edir və əlaqəli promoter fəallığı dərəcəsini proqnozlaşdırır. Bu ardıcıllıq, 600 bp uzunluğunda bir-isti-kodlanmış DNT ardıcıllığını təmsil edən (600, 4) formalı bir matris kimi göstərilir: A = [1, 0, 0, 0], C = [0, 1, 0, 0], G = [0, 0, 1, 0], T = [0, 0, 0, 1] və N = [0, 0, 0, 0]. MPRA fraqmentlərinin uzunluqları 88 bp-dən 600 bp-ə qədər dəyişdiyi üçün, 600 bp-dən qısa olan ardıcıllıqları maksimum uzunluğa çatana qədər N ilə doldurduq. Matrisin ortasındakı mövqelərə potensial meylliliyin qarşısını almaq üçün fraqment ardıcıllığının sağ və sol tərəflərinə əlavə edilən N əsaslarının sayı hər fraqment üçün vahid paylanmaya uyğun olaraq təsadüfi seçildi. Bundan əlavə, fraqmentlərin kənarlarında motivlərin yaranmasını nəzərə almaq üçün əsas plazmiddən adapter ardıcıllıqlarını əlavə etdik. Hər fraqmentin hər iki kənarına plazmid əsasına uyğun olan yeddi nukleotid ardıcıllığı daxil edildi. Xüsusilə, genom miqyaslı kitabxana üçün bu ardıcıllıqlar CAGTGAT və ACGACTG, hədəflənmiş kitabxana üçün isə CAGTGAT və CACGACG idi.
DL modelinin arxitekturası
Enformer-in ilk təbəqələrindən uyğunlaşdırılmış PARM arxitekturası, özünə diqqət toplama təbəqələri olan bir CNN-dən ibarətdir (Genişləndirilmiş Məlumat Şəkli 1a). Modellər hər bir hüceyrə növü üçün fərdi olaraq quruldu və hiperparametrlər proqnozlar ilə test dəsti ölçmələri arasındakı Pearson korrelyasiyasına görə optimallaşdırıldı. Birinci təbəqə, 7 nüvə ölçüsü və 125 filtr ölçüsü olan 1D konvolyusiya təbəqəsindən, ardınca bir qrup normallaşdırması, GeLU aktivasiya təbəqəsi və 1 nüvə ölçüsü və 125 filtr ölçüsü olan 1D konvolyusiyasından ibarətdir. İlk 1D konvolyusiya təbəqəsi ilə birinci təbəqənin çıxışı arasında sonrakı 125 filtr ölçülü diqqət təbəqəsini qidalandırmaq üçün bir keçid əlaqəsi əlavə edildi. Bu ilk blokdan sonra aşağıdakı ardıcıl təbəqələr 5 dəfə təkrarlandı: qrup normallaşdırması, GeLU aktivasiya təbəqəsi, keçid əlaqəsi ilə 1D konvolyusiya təbəqəsi (nüvə ölçüsü 7, doldurma 3, filtr ölçüsü 125), qrup normallaşdırması, GeLU aktivasiyası, 1D konvolyusiya (nüvə ölçüsü 1, filtr ölçüsü 125). Çıxış, keçid əlaqəsi ilə birləşdirildi və bir diqqət toplama təbəqəsinə verildi. Nəhayət, mövqe ölçüsü üzərində bir qlobal maksimum toplama təbəqəsi, ardınca 125 giriş ölçüsünü 1-ə endirən və GeLU aktivasiya təbəqəsindən keçən sıx bir təbəqə vardı. Model PyTorch (v.2.1.1) proqramında tətbiq edildi.
Modelin təlimi və qiymətləndirilməsi
Biz 30,607 promotoru 6 bərabər ölçülü qata böldük və üst-üstə düşən promotorların eyni qata təyin olunmasını təmin etdik. Müxtəlif məlumat qatları arasında ardıcıllıq fraqmentlərinin üst-üstə düşməməsini təmin etdik. Buna nail olmaq üçün, 2 kb + 600 bp məsafədə yaxın bir TSS olan hər hansı bir TSS eyni məlumat qatına təyin edildi. Bir qat son model qiymətləndirməsi üçün saxlanıldı, qalan beş qat isə çarpaz yoxlama üsulu ilə rotasiya edildi. Bu proses dörd qatın təlim üçün, bir qatın isə ən yaxşı hiperparametrləri müəyyən etmək üçün istifadəsini əhatə edirdi.
Biz promoter fəallığı balının qeyri-bərabər paylanmasını və ya heteroskedastik itkini nəzərə almaq üçün Puasson neqativ loqarifmik ehtimal itki funksiyasından istifadə etdik. Sinir şəbəkəsi cəmi 10 epoxa və 128-lik qrup ölçüsü ilə təlim edildi. Təlim prosesini optimallaşdırmaq üçün biz AdamW optimizatorundan istifadə etdik. Bu zaman, 2 × 10⁻⁸-dən başlayan və ilk 5,000 təlim qrupunda 10⁻⁴-ə çatana qədər xətti olaraq artan tədrici isinmə öyrənmə sürəti tətbiq edildi. Sonradan, öyrənmə sürəti son epoxaya qədər kosinus funksiyası ilə azaldıldı. Digər hiperparametrlər bunlardır: çəki azalması = 0.2, β₁ = 10⁻⁵ və β₂ = 10⁻⁵. Model bir NVIDIA Quadro RTX 6000 GPU-da təlim edildi.
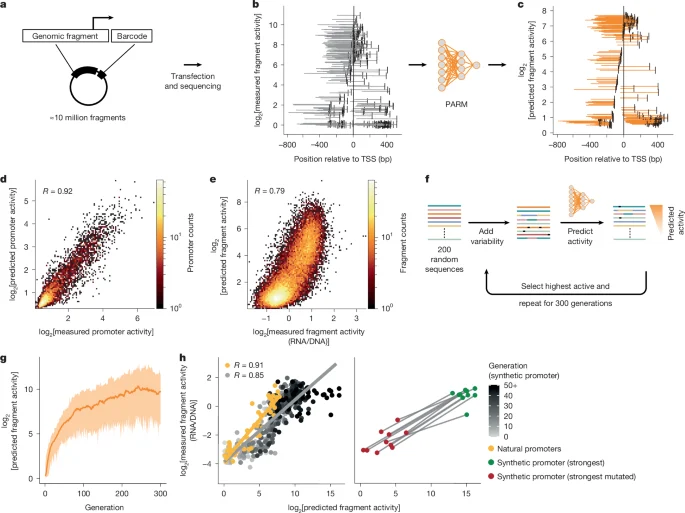
Aktiv sintetik promotorların yaradılması
PARM ilə sintetik insan promotorları yaratmaq üçün əvvəllər bildirilmiş genetik alqoritmdən istifadə etdik, lakin cüzi fərqlərlə. DEAP-ER (v.2.0.0, https://github.com/aabmets/deap-er) çərçivəsindən istifadə edərək, populyasiya ölçüsünü 200 olaraq təyin etdik, burada hər bir fərd 232 bp-lik təsadüfi yaradılmış bir ardıcıllıq idi. Mutasiya ehtimalını 0.8, iki nöqtəli krossover ehtimalını isə 0.5 olaraq təyin etdik və alqoritmi 300 nəsil boyunca işlətdik. Hər turda üç fərdin iştirak etdiyi standart turnir seçmə operatorundan (sel_tournament) istifadə etdik və qiymətləndirmə operatoru K562 modeli üçün PARM proqnozu idi. Genetik alqoritmin on müstəqil nümunəsini işlətdikdən sonra, bəzi sintetik və təbii ardıcıllıqları seçdik və onları reportyor kitabxanamıza klonlaşdırdıq.
Sintetik promotor kitabxanası
Genetik alqoritmin proqnozlarını təsdiqləmək üçün sintetik promotor kitabxanası qurduq. Fraqmentlər üçün aşağıdakı ardıcıllıqları seçdik: (1) Hər genetik alqoritm işləməsi üçün, 13 fərqli nəsildən (0-cı nəsildən başlayaraq 48-ci nəslə qədər hər dördüncü nəsil) ən az, orta və maksimum proqnozlaşdırılan aktiv fraqment olmaqla cəmi 390 (10 × 13 × 3) fraqment. (2) Hər genetik alqoritm işləməsindən (nəsildən asılı olmayaraq) ən aktiv proqnozlaşdırılan fraqment olmaqla cəmi on fraqment ki, biz bunları sintetik promotor (ən güclü) adlandırdıq. (3) Ən aktiv fraqmentlərin hər biri üçün, proqnozlaşdırılan aktivliyinin yalnız 20%-i qalana qədər həmin fraqmenti in silico mutasiya etdik. Bunlar da cəmi on fraqment idi və biz onları sintetik promotorlar (ən güclü mutasiyalı) adlandırdıq. (4) Təbii promotorlardan aşağıdan yuxarıya doğru aktivliklərə malik olan nəzarət ardıcıllıqları, o cümlədən K562 hüceyrələrində MPRA ilə ölçülmüş ən yüksək aktivliyə malik 15 promotor. Bu kitabxana yuxarıda təsvir edildiyi kimi klonlaşdırıldı. Fraqment aktivliyi dörd barkodun orta log₂[RNT/DNT] nisbətinin götürülməsi ilə hesablandı. Məlumatlar Əlavə Məlumat 1-də mövcuddur.
ISM təsdiqləmə kitabxanası
Hər bir fraqmentin aktivliyi, hər bir fərdi fraqmentin bağlı olduğu üç barkodun ortalaması alınaraq hesablandı. Etibarlı şəkildə ölçülmüş promotorlar və hüceyrə xətləri dəstini seçmək üçün avtokorrelyasiyalara əsaslanan sərt bir kəsmə dəyəri seçdik. Hər bir mövqe üçün bütün mutasiyaların təsirini bütün mutasiyaların ortalaması kimi hesabladıq. Sonra, birinci və sonuncu mövqelər istisna olmaqla, hər bir mövqenin sonrakı mövqe ilə Pearson korrelyasiyasını hesabladıq. Avtokorrelyasiyası 0.5-ə bərabər və ya daha yüksək olan promotor-hüceyrə xətti kombinasiyaları sonrakı təhlillər üçün saxlanıldı. Bu, etibarlı məlumatlara sahib olduğumuz 30 hüceyrə xətti-promotor kombinasiyasından ibarət bir dəstlə nəticələndi. Məlumatlar Əlavə Məlumat 2-də mövcuddur.
Motiv daxiletmə kitabxanası
Hər bir fraqmentin aktivliyi, hər bir fərdi fraqmentin bağlı olduğu beş barkodun ortalaması alınaraq hesablandı. Həmçinin, 0.01-lik bir psevdosay əlavə edildi. Bundan əlavə, hər bir fraqmentin ən azı 4 barkoda bağlı olmasını və pDNA (giriş) sayının >100 olmasını tələb etdik. Bu tələblər YY1 daxiletmələri olan 19, NFYA üçün 19, NRF1 üçün 19 və SP1 üçün 19 promotorla nəticələndi. Məlumatlar Əlavə Məlumat 3-də mövcuddur.
ZNF48 zülalının ekspressiyası
Tam uzunluqlu ZNF48 zülalını kodlayan kodon-optimallaşdırılmış gen bloku, Spodoptera frugiperda (Sf9) həşərat hüceyrələrində zülal istehsalı üçün əvvəllər təsvir edildiyi kimi Liqasiyadan Asılı Olmayan Klonlaşdırma vasitəsilə dəyişdirilmiş pFastBac vektoruna klonlaşdırıldı. Zülal amino-terminal 10×His-2×StrepII-etiketinə birləşdirildi. Bakmid DNT-si hazırlandı və Sf9 həşərat hüceyrələrində Invitrogen Bac-to-Bac metodu ilə P0 və P1 bakulovirus stokları istehsal edildi. Zülallar ESF921 mühitində (Expression Systems) kultivasiya edilən Sf9 həşərat hüceyrələrində ekspressiya edildi. Sf9 suspenziya hüceyrələri (hər ml-də 1 × 10⁶ hüceyrə) 28 °C-də 80 rpm-də çalxalayıcı inkubatorda kultivasiya edildi və P1 bakulovirus stoku ilə yoluxduruldu. Hüceyrələr infeksiyadan 3 gün sonra sentrifuqasiya (500g) ilə toplandı, maye azotda donduruldu və sonrakı emal üçün -20 °C-də saxlanıldı. Hüceyrə çöküntüsü əridildi və 25 mM Tris pH 8.0, 200 mM NaCl, 1 mM TCEP, 1 µM ZnCl₂ və 0.2% Triton X-100, 0.05% Tween-20 və 5% qliserol məhlulunda resuspensiya edildi. Hüceyrələr buz üzərində sonikasiya ilə lizis edildi və zibillər 4 °C-də 40 dəqiqə 53,340g-də sentrifuqasiya ilə uzaqlaşdırıldı.
Rekombinant ZNF48-in SF9 lizatından çökdürülməsi
30 bp-lik yabanı tip TMEM ardıcıllığını və ya onun mutasiyalı qarşılığını ehtiva edən sintetik oliqonukleotidlər, irəli zəncirdə 5′ biotinləmə ilə Integrated DNA Technologies şirkətindən sifariş edildi. Oliqonukleotidlər 95 °C-də 10 dəqiqə qızdırılaraq və tədricən otaq temperaturuna qədər soyudularaq oliqonukleotid hibridləşmə buferində (100 mM NaCl, 20 mM Tris-HCl pH 8.0 və 2 mM EDTA) hibridləşdirildi. Hər çökdürmə üçün 20 µl streptavidin sefaroza muncuq suspenziyası (GE Healthcare, GE17-5113-01) iki dəfə 0.1% NP-40 olan buz kimi soyuq PBS ilə, sonra isə bir dəfə DNT bağlama buferi (DBB) ilə yuyularaq tarazlaşdırıldı. Tarazlaşdırılmış muncuqlar daha sonra 500 pmol hibridləşdirilmiş oliqonukleotid qarışığı ilə 4 °C-də 30 dəqiqə yumşaq fırlanma ilə inkubasiya edildi. İnkubasiyadan sonra muncuqlar iki dəfə DBB ilə və bir dəfə zülal inkubasiya buferi (PIB) ilə yuyuldu. Hər çökdürmə üçün, rekombinant ZNF48 ifadə edən 1 və ya 2 mq SF9 hüceyrə lizatı oliqonukleotidlə bağlı muncuqlarla 4 °C-də 90 dəqiqə fırlanma ilə inkubasiya edildi. Muncuqlar sonradan beş dəfə PIB ilə və bir dəfə buz kimi soyuq PBS ilə yuyuldu. Bağlı zülallar muncuqların DTT olan 1× yükləmə nümunəsi buferində (Invitrogen, Bolt) resuspensiya edilməsi və 95 °C-də 10 dəqiqə qızdırılması ilə elüsiya edildi. Zibillər sentrifuqasiya ilə uzaqlaşdırıldı və nümunələr 4–15% TGX gradient gellərində (Bio-Rad) SDS-PAGE ilə ayrıldı. Zülallar iBlot 3.0 sistemi (Invitrogen) ilə 0.22 µm nitroselüloz membranına (Invitrogen, iBlot 3.0 transfer yığınları) köçürüldü. Membranlar otaq temperaturunda 30 dəqiqə ərzində 0.1% Tween-20 (PBS-T) olan PBS-də 5% yağsız süd ilə bloklandı. Membranlar 4 °C-də bir gecə dovşan poliklonal anti-ZNF48 antikoru (Sigma-Aldrich, HPA023806) ilə zondlandı. PBS-T ilə üç dəfə yuyulduqdan sonra, membranlar otaq temperaturunda 1 saat HRP-konjugasiyalı donuz anti-dovşan IgG IgG-HRP (Dako, P0399) ilə inkubasiya edildi. PBS-T ilə əlavə üç dəfə yuyulduqdan sonra, membranlar Fusion Fx görüntüləmə sistemi (Vilber) ilə görüntüləndi.
Kütlə spektrometriyası təhlili üçün hüceyrə lizatının hazırlanması
Xam nüvə ekstraktları əvvəllər təsvir edildiyi kimi hazırlandı. Hüceyrələr toplandı və fırladıldı, əldə edilən hüceyrə çöküntüsü iki dəfə buz kimi soyuq PBS ilə yuyuldu. Hüceyrə çöküntüsü 5 həcm A buferində resuspensiya edildi və buz üzərində 10 dəqiqə inkubasiya edildi. Hüceyrələr sentrifuqasiya ilə çökdürüldü və təzə hazırlanmış 2 həcm A+ buferində resuspensiya edildi. Hüceyrələr dounce homogenizasiyası ilə lizis edildi və xam nüvələr sentrifuqasiya ilə toplandı. Xam nüvələr buz kimi soyuq PBS ilə yuyuldu və təmiz xam nüvələr sentrifuqasiya ilə toplandı. Xam nüvələr C buferində resuspensiya edildi və 4 °C-də fırlanarkən 90 dəqiqə inkubasiya edildi. Sonra, nüvə lizatı sentrifuqasiya edildi və həll olunan nüvə fraksiyası toplandı. Əldə edilən nüvə ekstraktları alikvotlara bölündü, maye azotda donduruldu və -80 °C-də saxlanıldı.
DNT çökdürmələri
DNT oliqonukleotidləri irəli zəncirin 5′-biotinləməsi ilə Integrated DNA Technologies şirkətindən sifariş edildi. İstifadə olunan oliqonukleotidlərin ardıcıllıqları Əlavə Cədvəl 1-də verilir. Oliqonukleotidlər əvvəllər təsvir edildiyi kimi hibridləşdirildi. DNT çökdürmələri əvvəllər təsvir edildiyi kimi iki nüsxədə aparıldı. Hər reaksiya üçün 20 μl streptavidin-sefaroza muncuq suspenziyası (GE Healthcare) bir dəfə 1 ml 1% NP-40 olan PBS ilə və iki dəfə 1 ml DBB ilə yuyularaq tarazlaşdırıldı. Sonra, 500 pmol əvvəllər hibridləşdirilmiş DNT oliqonukleotidləri 600 μl yekun həcmdə DBB-də muncuqlarla inkubasiya edildi. Muncuqlar daha sonra iki dəfə DBB ilə və bir dəfə PIB ilə yuyuldu. Hər çökdürmə üçün 500 μg nüvə ekstraktı 600 μl yekun həcmdə PIB-də muncuqlarla inkubasiya edildi. İnkubasiyadan sonra muncuqlar 3 dəfə 1 ml PIB ilə və iki dəfə 1 ml PBS ilə yuyuldu. Artıq PBS şprislə çıxarıldı və muncuqlar dərhal 50 μl elüsiya buferində resuspensiya edildi və otaq temperaturunda silkələnərkən 20 dəqiqə inkubasiya edildi. Nümunələr daha sonra yodoasetamid ilə (50 mM yekun konsentrasiya) qaranlıqda silkələnərkən 10 dəqiqə inkubasiya edildi. Sonra, zülallar 0.25 μg tripsin ilə həzm edildi və otaq temperaturunda 2 saat silkələnərkən inkubasiya edildi. Nümunələr fırladıldı və supernatant toplandı. Muncuqlar bir daha 50 μl elüsiya buferi ilə yuyuldu, supernatant toplandı və əvvəllər toplanan supernatantın üzərinə əlavə edildi, qarışığa 0.1 μg tripsin əlavə edildi və bir gecə inkubasiya edildi. Növbəti gün nümunələr StageTips istifadə edilərək təmizləndi. StageTips-də dimetil etiketlənməsi əvvəllər təsvir edildiyi kimi aparıldı.
Kütlə spektrometriyası təhlili
Peptidlər Evosep One LC sisteminə (Evosep Biotechnology) qoşulmuş Orbitrap Exploris 480 kütlə spektrometrində LC-MS/MS ilə təhlil edildi. LC ayrılmasından əvvəl peptidlər 0.1% qarışqa turşusunda bərpa edildi və nümunənin 20%-i Evotip Pure (Evosep) ucluqlarına yükləndi. Peptidlər birbaşa sütuna elüsiya edildi və əvvəlcədən proqramlaşdırılmış "genişləndirilmiş metod" (88 dəqiqəlik qradiyent) ilə EV1137 (Evosep) sütununda EV1086 (Evosep) emitteri ilə birləşdirilmiş şəkildə ayrıldı. Nanosprey, sprey gərginliyi 1.7–2 kV-a ayarlanmış Easy-Spray NG İon Mənbəyi (Thermo) ilə əldə edildi. Exploris 480, 1 s ilə DDA dövr müddəti rejimində işlədildi. Tam MS skanları Orbitrap analizatorunda 375–1,500 m/z aralığında 200 m/z-də 60,000 rezolyusiya ilə toplandı. Defolt yük vəziyyəti 2+ olaraq təyin edildi, AGC hədəfi həm MS1, həm də ddMS2 üçün "standart" olaraq təyin edildi və hər iki skan növü üçün maksimum inyeksiya müddəti rejimi avtomatik olaraq təyin edildi. Monoizotopik pik müəyyənləşdirməsi peptid olaraq təyin edildi və dinamik istisna 20 s olaraq təyin edildi. ddMS2 üçün, 2+ ilə 6+ yük vəziyyətində olan və 5 × 10⁴ intensivlik həddi filtr kriteriyasına cavab verən prekursorlara 30% normallaşdırılmış HCD toqquşma enerjisi tətbiq edildi. Prekursorlar Kvadrupol analizatorunda 1.2 m/z izolyasiya pəncərəsi ilə izolyasiya edildi və MS2 spektrləri Orbitrap-da 15,000 rezolyusiyada əldə edildi.
Kütlə spektrometriyası məlumatlarının təhlili
Bütün xam kütlə spektrometriyası spektrləri Maxquant proqramı (v.2.4.9.0, Maks Plank Biokimya İnstitutu) ilə emal edildi və 2022-ci ildə yayımlanmış UniProt insan proteom məlumat bazasına qarşı axtarıldı. Müəyyən edilmiş zülallar ümumi çirkləndiricilər üçün süzgəcdən keçirildi. Dimetil etiketli nümunələr, daxili dimetilasiya 3plex metoduna əsaslanan dəyişdirilmiş iş axını ilə təhlil edildi. Kəmiyyət təyini üçün yüngül dimetil etiketli peptidlər (+28.031 Da) və ağır dimetil etiketli peptidlər (+32.056 Da) istifadə edildi. Aşağı bolluqda yenidən nümunə götürmə aktivləşdirildi. Yalnız dörd kanalda da kəmiyyəti müəyyən edilmiş zülallar sonrakı təhlil üçün istifadə edildi. Əhəmiyyətli zülalları müəyyən etmək üçün kənar statistika istifadə edildi. Həm irəli, həm də geri təcrübələr üçün bir kvartillərarası diapazona malik zülallar əhəmiyyətli hesab edildi.
ISM sınağı
ISM, bir istinad ardıcıllığının və maraq mövqeyində bütün mümkün tək mutasiyalara malik ardıcıllıqların promoter fəallığı proqnozunu əhatə edir. Əhəmiyyət balı, istinad ardıcıllığının balından tək bir mövqedəki bütün mümkün mutasiyaların orta balının çıxılması ilə hesablanır.
ICGC məlumatları
Beynəlxalq Xərçəng Genom Konsorsiumu (ICGC) məlumatları, IGCG məlumat portalının 28-ci buraxılışından əldə edildi. Tam genom sekvenslənməsi sekvensləmə strategiyası olaraq sorğulandı və hg19 istinad genomunda 5-ci xromosomda 1,295,152-dən 1,295,462 bp-ə qədər məlumatları olan bütün donorlar əldə edildi.
Enformer proqnozları
Enformer müqayisələri üçün variant təsirini əvvəllər təsvir edildiyi kimi proqnozlaşdırdıq. Müvafiq hüceyrə növləri üçün CAGE çıxışlarını hesabladıq və proqnozları loqarifmik çevirdik. Hər iki zənciri, SNP mövqeləri ətrafındakı üç qonşu qutunu və müvafiq hüceyrə xətləri üçün mövcud olan bütün CAGE izlərini ortaladıq. Təsdiqləmə kitabxanamızdakı bəzi hüceyrə xətləri Enformer-də olmadığı üçün AGS hüceyrə xəttini sınaqdan keçirmək üçün GSS, MKN45, AZ521 və MKN1; U2OS üçün HS-Os-1; HCT116 üçün CACO-2 və COLO-320; LNCaP üçün isə DU145 istifadə etdik.
Borzoi proqnozları
Borzoi proqnozları üçün variant təsirini əvvəllər təsvir edildiyi kimi proqnozlaşdırdıq. Yuxarıda Enformer-də CAGE istifadə edərək təsvir edilən eyni proseduru izlədik. Çatışmayan hüceyrə xətləri üçün AGS hüceyrə xəttini sınaqdan keçirmək üçün GSS, MKN45, AZ521 və MKN1; U2OS üçün HS-Os-1; LNCaP üçün DU145; HCT116 üçün isə CACO-2 və COLO-320 istifadə etdik.
cis-təsirli eQTL-lərin təsir istiqamətinin proqnozu
PARM, Enformer və Borzoi modellərinin proqnozlaşdırılan ekspressiya təsirinin dəqiqliyini qiymətləndirmək üçün, proqnozlaşdırılan təsir istiqamətini dəqiqləşdirilmiş GTEx v8 cis-təsirli eQTL-lərin ölçülmüş beta işarəsi ilə müqayisə etdik. Bu dəstdən, SuSIE tərəfindən müəyyən edilmiş kimi posterior səbəb ehtimalı (PIP) ≥0.9 olan etibarlı bir dəstdəki bütün variantları götürərək tam qandan bütün dəqiqləşdirilmiş ehtimal olunan səbəbkar cis-təsirli eQTL-ləri seçdik. Biz yalnız dəqiqləşdirilmiş tək nukleotid variantlarını (SNV) nəzərə aldıq, çünki insersiya/delesiya təsirlərini modelləşdirmək daha çətindir və SNV-lərin proqnozlaşdırılan təsir ölçülərindən əhəmiyyətli dərəcədə fərqlənir. Təsir istiqamətindəki uyğunluq aşağıdakı kimi müəyyən edilir:
İstiqamətdə Uyğunluq = (ΣNeQTLsi=1 1(variant təsiriᵢ ⋅ βᵢ > 0)) / NeQTLs
Burada NeQTLs sıfırdan fərqli təsiri olan eQTL-lərin ümumi sayını (1,836) və β isə GTEx-də təxmin edilən müşahidə olunan eQTL təsirini göstərir. Əsas müqayisə kimi, NumPy-nin random.uniform tətbiqi ilə -1-dən +1-ə qədər təyin edilmiş vahid paylanmadan təsadüfi nümunə götürərək təsadüfi proqnozlar üçün istiqamət uyğunluğunu hesabladıq.
Motiv skan edilməsi və RS identifikasiyası
Hocomoco (v.11) insan məlumat bazasından istifadə edərək, (hüceyrə tipinə məxsus) PARM modellərindən biri tərəfindən yaradılan 30,607 promotorun əhəmiyyət ballarını skan etdik. Promotor əhəmiyyət ballarını ehtiva edən bu 30,607 vektorun hər biri, Hocomoco məlumat bazasından hər bir informasiya məzmunu (IC) motivi ilə 1 addımla skan edildi. Hər mövqedə, motivin IC-si ilə hər bir mövqedəki əhəmiyyət balı arasındakı Pearson korrelyasiya əmsalını hesabladıq. Bundan əlavə, IC ilə vurulmuş təsir ballarının ortalamasını hesabladıq. Biz yalnız Pearson korrelyasiya əmsalı |0.75|-ə bərabər və ya daha yüksək olan nəticələri saxladıq. Əhəmiyyət balının aşağı olduğu, lakin korrelyasiyanın 0.75-dən yüksək olduğu bölgələrdəki nəticələrdən qaçmaq üçün (bu, xüsusilə qısa və yüksək məzmunlu motivlər üçün baş verirdi), motivlərin dəyişən uzunluqlarını və IC-ni nəzərə alan ikinci bir kəsmə tətbiq etdik. Bir RS-nin müəyyən edilməsi üçün həm korrelyasiya, həm də təsir balı kəsimlərindən keçməsi tələb olunurdu. Ölçülmüş məlumatlarda RS-lərin identifikasiyası üçün eyni şəkildə korrelyasiya hesablandı, lakin təsir balı kəsimi müvafiq modeldən idxal edildi, Z-balına çevrildi və sonra ölçülmüş məlumatlara tətbiq edildi.
RS paylanma müqayisələri
RS paylanması ilə promotorlar arasında spesifik TF-lərin ümumi motiv paylanması arasındakı fərqi müəyyən etmək üçün, iki empirik paylanma arasında hər hansı bir fərqi (yer, miqyas və ya forma) müəyyən etmək üçün istifadə olunan Kolmoqorov-Smirnov testindən istifadə edildi. RS paylanmaları arasında rejimdə bir dəyişikliyi xüsusi olaraq test etmək üçün (aktivləşdirici və repressiv RS-lərin müqayisəsində olduğu kimi) bir permutasiya testindən istifadə etdik. Aktiv və ya repressiv sinfə təyinat qarışdırıldı, bundan sonra repressiv RS paylanmasının rejimləri ilə aktivləşdirici RS paylanmasının rejimləri arasındakı fərqin boş paylanması müəyyən edildi. Bu boş paylanma empirik bir P dəyərini hesablamaq üçün istifadə edildi.
Naməlum motivlərin müəyyən edilməsi
Heç bir məlum motivə uyğun gəlməyən RS-ləri müəyyən etmək üçün, bütün əhəmiyyət ballarının aktivləşdiricilər üçün Əhəmiyyət balı₉₅%-dən bərabər və ya yüksək, yaxud repressivlər üçün Əhəmiyyət balı₅%-dən bərabər və ya aşağı olduğu ən azı beş nukleotidlik sahələri skan etdik. Maksimum iki nukleotidlə ayrılmış bir neçə ardıcıl RS varsa, onlar tək bir RS kimi qəbul edildi. Sonra, tam RS bölgəsinin nukleotid əhəmiyyəti Hocomoco (v.11) insan və ya Jaspar2024 CORE onurğalılar qeyri-artıq məlumat bazasındakı bütün motivlərlə korrelyasiya edildi. Heç bir məlumat bazasında R ≥ 0.75 olan bir uyğunluq yoxdursa, o, "naməlum motiv" kimi qeyd edildi.
Motiv ailələri əldə etmək üçün motivlərin klasterləşdirilməsi
Hocomoco (v.11) insan məlumat bazası və naməlum motivlər üçün motivləri ayrıca klasterləşdirmək üçün əvvəllər təsvir edilmiş metodları izlədik. Mövqe tezlik matrislərinin cütlüklü oxşarlıq balları, minimum 6 bp üst-üstə düşmə və hizalanmaların qiymətləndirilməsi üçün məsafə ölçüsü kimi Pearson korrelyasiyası ilə TOMTOM (v5.4.1) istifadə edilərək hesablandı. Sonradan, bu balları 1 × 10⁻⁸ psevdosay əlavə etdikdən sonra loqarifmik çevrilmə ilə E dəyərlərinə çevirdik. İyerarxik klasterləşdirmə daha sonra tam əlaqə və məsafə ölçüsü kimi Pearson korrelyasiyası ilə aparıldı. Ağac 0.8 hündürlüyündə kəsildi və kəsim daxilindəki bütün motivlər eyni motiv ailəsindən hesab edildi.
RS-in TF ekspressiyası ilə uyğunlaşdırılması
Sonrakı təhlil üçün yalnız ifadə olunan TF-lərlə əlaqəli RS-ləri daxil etdik. İfadə statusu İnsan Zülal Atlasından (v.23.0) əldə edilən məlumatlar əsasında müəyyən edildi. ln(TPM+1) ≥ 1 olan TF-lərlə əlaqəli RS-lər sonrakı təhlillər üçün seçildi.
Konsensus motiv
Naməlum motivlər klasterində konsensus motivini qurmaq üçün nümayəndə motiv kimi ən aşağı E dəyərinə malik motivi seçdik. Klasterdəki hər bir motiv üçün, onun mövqe tezlik matrisi TOMTOM tərəfindən bildirilən optimal ofset və oriyentasiya nəzərə alınmaqla nümayəndə motivə əlavə edildi. Hər mövqe daha sonra həmin mövqeyə əlavə edilmiş motivlərin sayına görə normallaşdırıldı. Konsensus motivində bir mövqeyə klasterdəki motivlərin 30%-dən azı töhfə veribsə, həmin mövqe silindi.
Ardıcıllıq skan edilməsi
TSS-nin 250 bp yuxarı axınından 50 bp aşağı axınına qədər olan bölgəni əhatə edən bütün promotor ardıcıllıqlarının sadə bir skanını FIMO (v5.4.1) ilə defolt arqumentlər və Hocomoco (v.11) insan məlumat bazası ilə həyata keçirdik.
RS daxiletmə təhlili
YY1, SP1, NRF1 və NFYA motivlərinin bütün promotorlara in silico daxil edilməsini həyata keçirdik. Bu TF-lərin hər biri üçün üç variant daxil etdik: konsensus ardıcıllığı və neqativ nəzarət kimi iki mutasiyalı versiya. Bənzər motiv uzunluğu aralıqlarında işləmək üçün Hocomoco (v.11) tərəfindən müəyyən edilmiş konsensus ardıcıllığını qısaltdıq. Ardıcıllıqlar mətnin orijinal hissəsində ətraflı şəkildə təsvir edilmişdir. Motivlər, TSS-nin 160 bp yuxarı axınından 63 bp aşağı axınına qədər olan 15 mövqeyə, 14 nukleotidlik sabit addımla daxil edildi.
Endogen promoter fəallığı
K562 hüceyrələrində endogen promoter fəallığı təxminini əldə etmək üçün aşağıdakı məlumat dəstlərini birləşdirdik: GRO-cap, csRNA, NetCage və Stripe-Seq. Xam faylları (fastq faylları) əldə etdik və hər oxunuşun sonundan 20 nukleotidi kəsmək üçün FASTX Toolkit (v.0.0.14) istifadə etdik. Sonra, oxunuşları insan rDNT-sinə qarşı hizalamaq üçün Bowtie2 (v.2.5.1) istifadə etdik. rDNT-yə hizalanmayan oxunuşlar daha sonra hg19 istinad genomuna xəritələndi. BamCoverage (v.3.5.2) istifadə edərək bigwig faylları yaratdıq. Bu bigwig faylları daha sonra 30,607 TSS dəstimizlə üst-üstə qoyuldu. Nəhayət, əsas komponent təhlili apardıq və variasiyanın 94.19%-ni izah edən ilk komponenti endogen promoter fəallığı göstəricisi kimi istifadə etdik.
Ölçülmüş diferensial promoter fəallığı təhlili
Fərqli şərtlər üçün ölçülmüş məlumatlarda diferensial promoter fəallığı təhlilinə daxil olmaq üçün əvvəlcə promoter fəallığı dəyərlərinə LOESS korreksiyası tətbiq etdik, sonra şərtlərin promoter fəallığı üzərindəki təsirlərini qiymətləndirmək üçün Uilkokson işarəli ranq testi apardıq. Xam P dəyərləri FDR-ə düzəldildi. Düzəldilmiş P dəyərləri 0.05-dən aşağı olan promotorlar üçün log₂[qat dəyişikliyi] > 0.5 olanları yuxarı tənzimlənmiş, log₂[qat dəyişikliyi] < -0.5 olanları isə aşağı tənzimlənmiş hesab etdik.
Hesabat xülasəsi
Tədqiqat dizaynı haqqında əlavə məlumat bu məqalə ilə əlaqəli Nature Portfolio Hesabat Xülasəsində mövcuddur.
Mənbəyə görə, bu xəbər vacib bir mövzudur. Daha çox xəbər üçün səhifəmizi izləyin.Fikirlərinizi bölüşün!
Oxucu Şərhləri
Hələlik heç bir şərh yazılmayıb. İlk şərhi siz yazın!
Şərh Yaz